H.O. Baral, Tübingen, 2004
| introduction | general remarks, vital taxonomy |
| morphology | apothecia (with images),asci and ascospores (with images), paraphyses (incl. pigments), excipulum & hairs (with images)
ascospore germination, anamorphs, substrate colonization |
| ecology | water supply, phenology, tolerancehost specifity, plant communitiesgeographic distribution (with map of Central Europe) |
| taxonomy | key to the seven European and N-American taxaphylogeny within Sarcoscyphaaccepted and excluded taxa, references |
Introduction
back to contentsGeneral remarks
Sarcoscypha, the type genus of the family Sarcoscyphaceae (Pezizales, operculate discomycetes, Ascomycotina), is well known in Great Britain as “scarlet-cups” by its bright scarlet- or purple-red discs, “perhaps the most beautiful of British ascomycetes” (Dennis 1978). The genus is widespread in north temperate and boreal regions, but also occurs in (sub)tropical areas and in the southern hemisphere. To the present knowledge, about 18 species can tentatively be accepted world-wide, about 6 of which occur within Europe and N-America. Many appear to be endemic to volcanic islands in the subtropics. Though quite a high number of species (about 60) have ever been combined in the genus Sarcoscypha, many of these remained virtually unknown as they were often only collected a single time, and especially the old descriptions are very inadequate. Often redescriptions of the type material, if any exists, are lacking. Many of the taxa have later been found to belong in other genera of the Sarcoscyphaceae, or were referred to other families of the Pezizales, some even to the Helotiales.
Unlike most Pezizales, the members of the Sarcoscyphaceae grow as saprophytes on dead woody plant material (rarely woody fruits), in the case of Sarcoscypha on mostly damp, mossy branches of broad-leaved, very rarely coniferous trees, that were fallen to the ground. Most species of Sarcoscypha fruit in winter and early spring, and are often already mature when still covered by snow.
back to contentsVital taxonomy
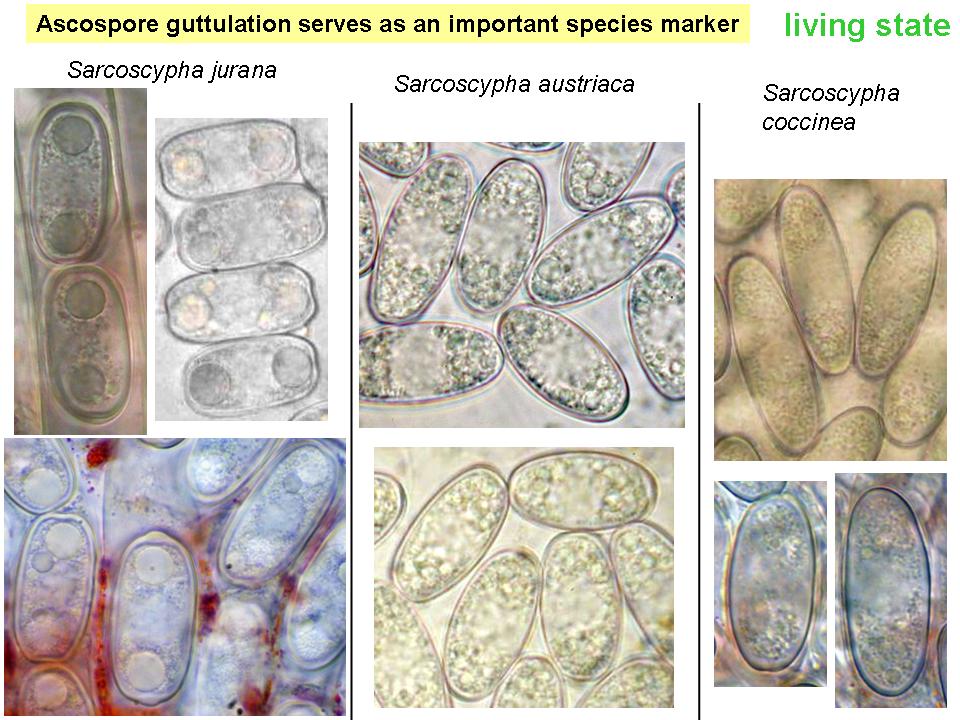
Although microscopical differences observed in living specimens has indicated since about 1900 the existence of several, macroscopically indistinguishable taxa within the temperate to alpine-boreal zone of the northern hemisphere (Europe and North America), only one large-cupped species of Sarcoscypha, S. coccinea, has currently been recognized in this large area during many later decades. The diagnostic microscopical characters supporting existence of different taxa mainly concern (1) striking differences in the guttule (lipid) pattern of living ascospores, first observed by Boudier (1903), (2) a very different germination behaviour of the ascospores among collections (with or without forming conidia), first observed by Rosinski (1953, corresponding case reported by Paden 1974 in Phillipsia), and (3) differently shaped spore ends. Strong differences in conidial size were later observed in pure culture, and DNA sequences supported existence of different taxa.
Baral (1984, Central Europe), Harrington (1990, USA), Butterfill & Spooner (1995, Great Britain), Pidlich-Aigner (1999, Austria), Öpik et al. (2000, Estland), G. & Y. van Duuren (2003, 2004, Netherlands), Matočec & Kušan (2007, Croatia) and B. & O. Perić (2007) confirmed the observations of Boudier and Rosinski, and showed that totally four different large-cupped species exist in Europe and N-America (S. austriaca, S. coccinea, S. dudleyi, S. jurana, apothecial diameter 20-90 mm), three of them in Europe. Both geographical distribution and host specifity differ severely among these species. Two further species deviating by smaller apothecia and also smaller ascospores exist in that area. One is perhaps endemic to the Macaronesian islands (S. macaronesica, unclear forms intermediate to S. coccinea appear to be characteristic of the eumediterranean belt). The other species occurs in atlantic and continental N-America east of the Rocky Mts. (S. occidentalis).
Both lipid pattern and germination behaviour are vital characters, which means that they are difficult or impossible to observe in dead herbarium material: (1) ascospore guttulation severely alters in dead spores by fusion of the oil drops, and (2) presence of germinated ascospores in fruit-bodies strongly depends on the development stage of the preserved collection. Fresh samples can be kept moist until spores germinate abundantly. If dried samples show fused oil drops and do not contain germinated spores, the differences between the species bescome quite obscure. As a consequence, herbarium taxonomists like Le Gal (1941) were unable to confirm the differences as reported by Boudier and Rosinski.
Harrington (1990: 436) wrote:“The importance of fresh material for species diagnosis, especially for noting ascospore guttulation, cannot be overstated. Although I had examined material (dried herbarium specimens) from western North America I was not prepared to recognize that group as a species distinct from the two, large eastern North American species until I saw fresh (living) material.”Sarcoscypha represents one of the numerous examples of fungal genera in which a sound taxonomy is only achieved on the basis of vital macro- and especially microscopical characters gained from the study of fresh collections (“vital taxonomy”).
back to contents
Morphology
back to contentsApothecia
The size of the apothecia ranges from about 1 to 8 cm in diam. Macroscopically the species of Sarcoscypha look very similar, therefore are hardly distinguishable. No clear difference in colour was found (except for the consistently white S. vassiljevae). However, recent observations indicate that S. coccinea might differ from both S. austriaca and S. jurana by a more hairy to crenulate apothecial margin. genetic variants with yellow or white hymenia rarely occur due to lacking colour components (see under paraphyses).
 Sarcoscypha austriaca (S-Germany, Black Forest, 4.IV.1993, del. O. Baral) Sarcoscypha austriaca (S-Germany, Black Forest, 4.IV.1993, del. O. Baral) |

Sarcoscypha jurana (Luxembourg, Fielsmillen, Tilia, 16.III.2004, G. Marson, phot. HB) |
 Sarcoscyphajurana (S-Germany, Swabian Alb, Tilia, 22.XI.1978, phot. HB) Sarcoscyphajurana (S-Germany, Swabian Alb, Tilia, 22.XI.1978, phot. HB) |
|
 Sarcoscypha austriaca (S-Germany, Füssen, Acer pseudoplatanus, 5.IV.1982, phot. A. Bollmann) Sarcoscypha austriaca (S-Germany, Füssen, Acer pseudoplatanus, 5.IV.1982, phot. A. Bollmann) |

Sarcoscypha coccinea (?macaronesica) (S-Italy, Pyrus pyrainus, 16.XII.1982, phot. C. Lavorato) |
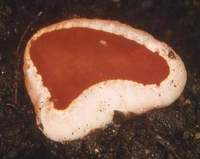 Sarcoscypha macaronesica (Tenerife, ?Lauraceae, 18.I.1996, phot. H. Grünwald) Sarcoscypha macaronesica (Tenerife, ?Lauraceae, 18.I.1996, phot. H. Grünwald) |
|
 Sarcoscypha occidentalis (USA, Ithaca NY, Rosales?, 20.IX.1983, phot. T.R. Lohmeyer) Sarcoscypha occidentalis (USA, Ithaca NY, Rosales?, 20.IX.1983, phot. T.R. Lohmeyer) |
 Sarcoscypha coccinea (?macaronesica) (S France, Quercus ilex, 16.I.2002, phot. G. Garcia) Sarcoscypha coccinea (?macaronesica) (S France, Quercus ilex, 16.I.2002, phot. G. Garcia) |
 Sarcoscypha occidentalis ( ?or S. javensis) (?USA, phot. P. Kaminski, from internet) Sarcoscypha occidentalis ( ?or S. javensis) (?USA, phot. P. Kaminski, from internet) |
|

Sarcoscypha coccinea (N Germany, Wolfenbüttel, Fagus, 1.III.2004, phot. U. Kozik) |
|||

Sarcoscypha austriaca (S Germany, Ummendorf, Alnus, 22.III.2004, K. Keck, phot. HB) |

Sarcoscypha coccinea (France, Villiers, Crataegus, 14.II.2004, J.P. Priou, phot.JPP) |

Sarcoscypha jurana, Luxembourg, Fielsmillen, Tilia, 16.III.2004, Guy Marson, phot. HB Right: Sarcoscypha coccinea (N Germany, Wolfenbüttel, Fagus, 1.III.2004, phot. U. Kozik) |
 |
Asci
Ascus
base.The asci of Sarcoscypha and also other
Sarcoscyphaceae, to the present knowledge, consistently arise from simpe septa.
This is in contrast to the other families of the Pezizales, in which the asci
mostly arise from croziers.
Ascus
apex. See under “Ascus Apex
Morphology and Iodine Reaction in Ascomycetes” (in preparation).
Ascospores
Shape. In S.
austriaca and S. jurana the
truncate, often indented (notched) poles have the shape of a saddle, so that a
turn of the spore for 90° along its longitudinal axis results in a
hemispherical to obtuse aspect of both poles. Therefore only about 50% of the
spores in a preparation show truncate poles (those seen in front view). This
fact is frequently mistaken as a variability in spore shape. If the spores are
dead, the degree of truncation or indentation is enhanced because of the
absence of an internal cell turgor. The function of this peculiar spore shape
is unknown; germination appears never to take place at the saddle but rather at
the edges (“shoulders”) or laterally.
|
Mature ascospores of Sarcoscypha, living state |
|
|
|
Sarcoscypha austriaca (Salix, Tübingen, 31.3.2003, leg. H.O. Baral) Sarcoscypha austriaca (Tegernsee, Mangfall, 4.2006, leg. ?)
|
Sarcoscypha jurana (Tilia, Oberpfalz, Dietfurt, 6.3.2012, leg. C. Wolf)
Sarcoscypha coccinea (Fraxinus, Lauffen, 4.2.2009, leg. H. Weckermann) |
|
|
|
|
|
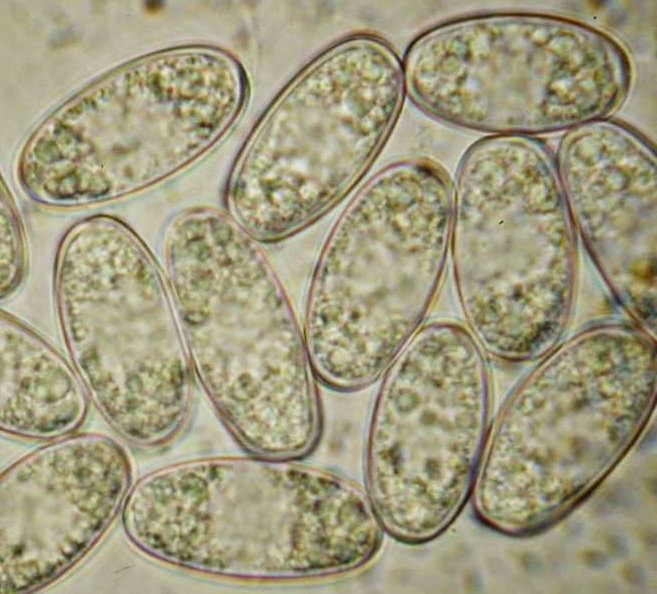
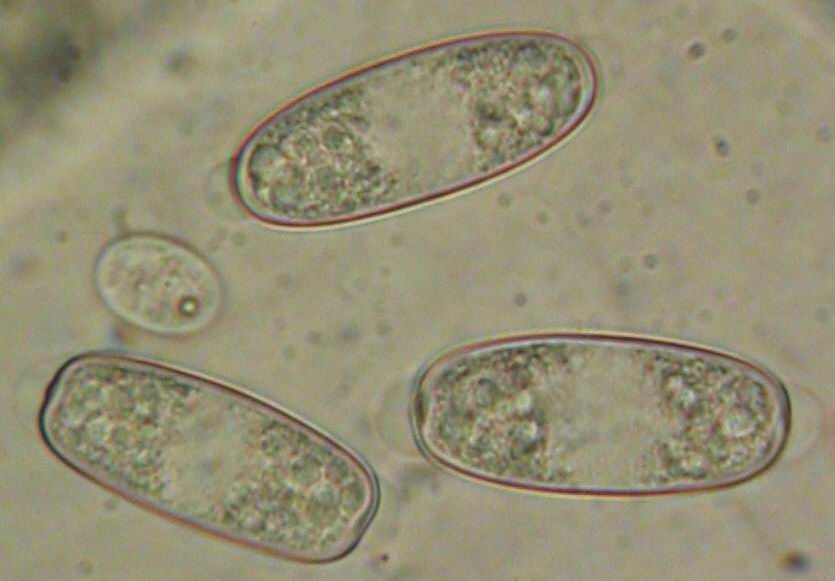
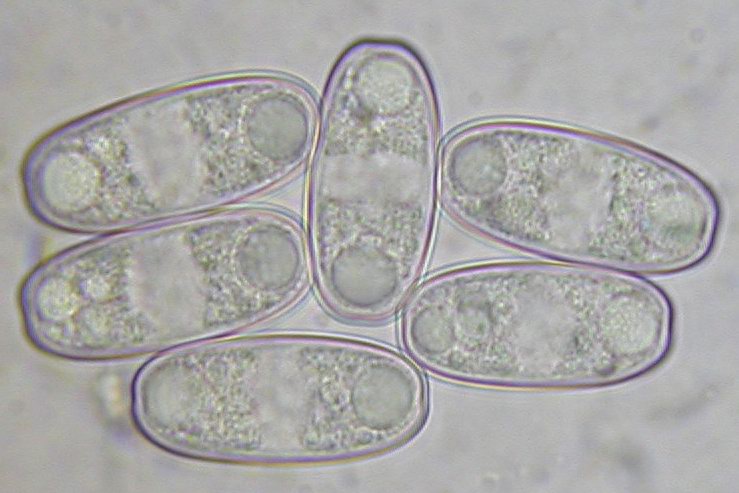
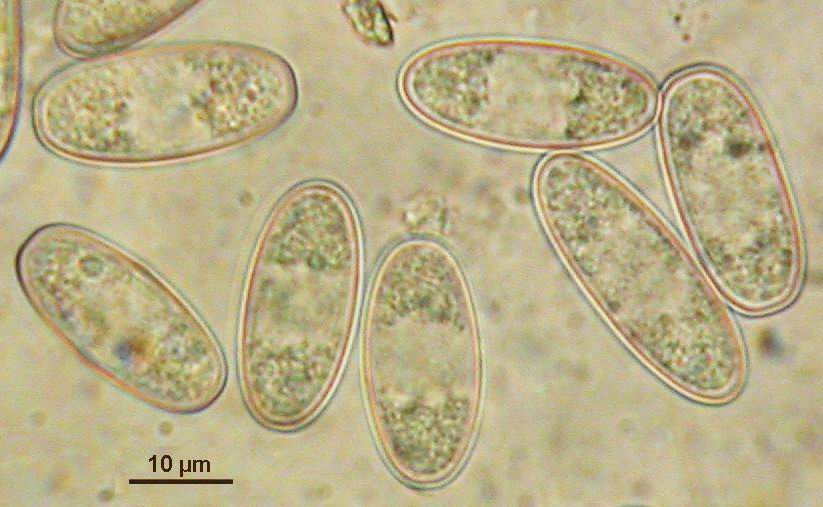
Mature ascospores of Sarcoscypha, living vs. dead state
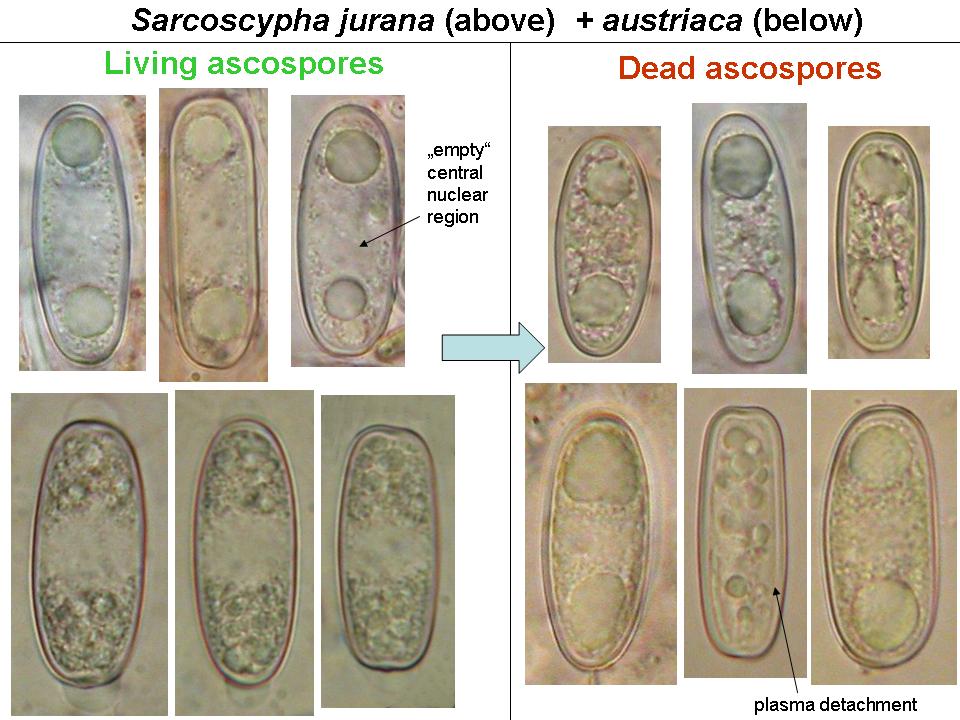
The strong difference in guttule pattern in the living spores between S. jurana and S. austriaca (left column) completely disappears in dead spores due to coalescence of the oil drops, the spore interior thereby looking quite variable (right column).
Therefore, herbarium material older than about 5-10 years is not easy to identify.
Contents.Characteristic
of the genus and perhaps the whole family is the presence of numerous
refractive minute oil drops (lipid bodies, LBs) among some larger LBs within the
living mature spores. A central area is free of LBs and contains 32 small
nuclei (?16 in S. occidentalis). Within Sarcoscypha only the
larger LBs are of taxonomic importance for species delimitation. While the
large and medium-sized LBs (characteristic of S. jurana and S. austriaca),
but also the minute LBs are rapidly seen in living spores, the relatively small
LBs of S. coccinea (see photo) and S. macaronesica are easily overlooked
among the minute ones. In dead spores the LBs are difficult to see because the
cytoplasma gets refractive when observed in a water mount. Mounting in KOH
makes the LBs reappear in full strength. However, coalescence of both larger
and smaller LBs takes place in many of the dead spores causing a very variable
guttule pattern which makes species recognition extremely difficult (see table
above). The specific guttule pattern is often already destroyed when the spore
powder got dry for only a few hours. On the other hand, some spores may still
show the unaltered guttule pattern even in very old herbarium material, helping
in recognizing the species.
|
S. |

Mucilaginous envelope (sheath). In several of the species the mature ascospores
are surrounded by a delicate envelope. Within the living mature asci the
envelope is normally strongly compressed and of higher refractivity compared to
those spores within dead asci or being ejected. During immature stages,
however, an envelope is probably present in all taxa. The envelope is a very
delicate, evanescent structure which disappears in a watery environment some
period of time after ejection, and which is rather impossible to detect in
herbarium material. However, sheaths can well be seen in water mounts made from
spore prints several years after drying (Peric 2008). The envelope does not stain
in aqueous cresyl blue or Toluidin blue, while the spore wall surface stains
deep lilac in these reagents (only remnants of dead epiplasma around the
envelope stain violet).
The shape of the envelope
is rather constant within a species and serves as a valuable characteristic for
species delimitation. However, the envelope may be absent depending on the
geographical origin. S. austriaca usually has polar cap-like envelopes,
but sometimes no envelope at all. S. coccinea in Europe is completely
devoid of an envelope while in N-America an envelope around the whole spore
similar as in S. jurana is present
(Harrington 1990).